
В ожидании Гого. Рыба, которая была готова пойти
В издательстве «Альпина нон-фикшн» вышла книга «Как живые: Двуногие змеи, акулы-зомби и другие исчезнувшие животные» геолога Андрея Журавлева. Автор рассказывает о необычных ископаемых животных, позволяющих понять эволюцию позвоночных: от мелких организмов, плававших в морях почти 520 млн лет назад, до гигантских ящериц, чьи шаги сотрясали почву всего чуть более 40 000 лет назад. Публикуем главу, посвященную древним мясистолопастным рыбам, которые в чем-то уже уподобились первым наземным позвоночным.
История латимерии, поимка которой в 1938 г. на юго-западе Индийского океана потрясла научный мир не меньше, чем если бы где-нибудь в дебрях Конго увидели живого игуанодона или птеродактиля, известна многим. Об этом удивительном событии можно узнать, например, из трех замечательных книг. Равноправные авторы этой сенсации — куратор скромного музея в Ист-Лондоне (ЮАР) Марджори Куртене-Латимер, любознательный и отважный капитан траулера Хендрик Госен и преподаватель химии из Университетcкого колледжа Родса в Грейамстауне (но в душе ихтиолог) Джон Смит. Именно он впоследствии станет автором научного описания удивительной рыбы, заплывшей в наше время прямо из мезозойских вод, и напишет книгу, переведенную на многие языки, в том числе на русский. Статья так и называлась — «Живая рыба мезозойского образа» — и предварялась крылатой фразой античного ученого Плиния Старшего: «Из Африки всегда что-то новое». Мисс Латимер была заслуженно увековечена в родовом имени самой удивительной рыбы на свете, а речка Чалумна, к устью которой вблизи портового городка Ист-Лондона заплыл тот самый целакант, чтобы угодить в трал капитана Госена, — в видовом (Latimeria chalumnae).
Дело в том, что все родственники латимерии — целаканты, одна из многочисленных в прошлом групп мясистолопастных рыб, — приказали долго жить еще до мел-палеогенового массового вымирания (около 70 млн лет назад). Они полностью исчезли из ископаемой летописи. И вдруг — настоящий целакант! Палеонтологам пришлось даже придумать новый термин — «виды Лазаря». Так называют виды, которые вроде бы вымерли, но вдруг оказались живы. (Латимерия в этом смысле отнюдь не исключение. В современных морях встречаются, например, строматопороидные и хететидные обызвествленные губки: они тоже как бы исчезли в мезозойскую эру.)
Ныне целаканты обитают на относительно больших глубинах (150–600 м) в районе Мозамбикского пролива и в противоположном «углу» Индийского океана — внутренних морях Индонезии (Latimeria menadoensis). Причем между островами Сулавеси и Новая Гвинея образовалось две популяции, особи которых не скрещиваются между собой более 10 млн лет — их развели морские течения. Ископаемых целакантов насчитывается под сотню видов и около 30 родов. Лучше всего им жилось в раннетриасовую эпоху: в водоемах, почти освободившихся от других палеозойских рыб, эти удивительные создания быстро заполонили все пространство. Таких приспособленцев палеонтологи называют «виды бедствия» (англ. disaster species). Как в известной поговорке: что русскому здорово, немцу смерть. Возможно, именно это умение получать выгоду в самой невыгодной ситуации и сохранило латимерию до наших дней.

Хотя многое о мясистолопастных мы знаем по ископаемым остаткам, изучение латимерии подтвердило, что основой осевого скелета у таких рыб служит хорда, тел позвонков нет, но есть мясистые и длинные парные грудные и брюшные плавники. Анальный и второй спинной плавники устроены точно так же, как парные. Это сходство еще раз подчеркивает тот факт, что и парные, и непарные плавники лишь чуть-чуть различаются на генном уровне.
Грудные и брюшные лопасти поддерживаются мощным скелетом из удлиненных костей. В них можно распознать те же элементы, что и в наших конечностях: плечевую, локтевую и лучевую кости на «руках»; бедренную, больше- и малоберцовую на «ногах». Есть также плечевой и тазовый пояса скелета. Эта конструкция в сочетании с мощными мышцами позволяет грудным плавникам свободно вращаться, принимая практически любое положение. Причем «походка» латимерии — как у четвероногих: сначала гребок одновременно совершают левая передняя и правая задняя конечности, затем движение повторяет другая пара. Анальный и спинной плавники при этом синхронно отклоняются то вправо, то влево, как единое рулевое весло, не давая телу вихлять из стороны в сторону. Трехраздельная (как бы с модным хвостиком-«косичкой» посередине) хвостовая лопасть остается неподвижной и включается в работу, лишь когда рыбе нужно ускориться.
Тело покрыто эллипсовидной крупной (до 15 см длиной у ископаемых форм) чешуей особого (эласмоидного) типа. У африканской латимерии она очень яркая — цвета индиго. Чешуя состоит из тонкого внешнего костного и гораздо более массивного внутреннего коллагенового слоев. Коллагеновая подложка — эласмодин — сложена многочисленными волокнистыми пластинами, причем волокна в соседних пластинах упакованы крест-накрест. На первый взгляд такая чешуя кажется слабой и ненадежной. Это впечатление обманчивое: рыбья шкура настолько крепкая, что на Коморских островах ее используют вместо наждака. Латимерии чешуя, конечно, нужна не в этом качестве: далеко налегающие друг на друга упругие коллагеновые пластины надежно защищают тело от укусов акул равного и чуть большего размера. Шкура выдерживает захват острыми зубами с силой до 25 Н. От больших акул спасает глубина, где целаканты и выжили. Костная поверхность чешуи, в свою очередь, несет многочисленные ребрышки и шипики, которые при движении рыбы ориентированы вдоль встречного потока и тем самым уменьшают лобовое сопротивление среды.
Крючьевидные острые конические зубы и/или мелкие зубчики образуют две дуги: снизу — на зубных и венечных костях, сверху — на челюстных, нёбнокрыловидных и сошниках. Зубы покрыты эмалью, а основу образует складчатый дентин (плицидентин) — так же как у древних четвероногих позвоночных. (Раньше по таким зубам выделялась большая группа земноводных — лабиринтодонты.)
Жаберные дуги несут костяные зубцы вместо мягких жаберных тычинок. Есть спиральный клапан в кишечнике и артериальный конус в сердце, как у живых и ископаемых представителей древних эволюционных ветвей костных и хрящевых рыб. Осморегуляция (выравнивание давления между морской водой и внутренней средой) осуществляется удержанием в крови мочевины и триметиламин-N-оксида, а также с помощью ректальной железы, выделяющей избыточную соль, — опять же как у хрящевых. Присутствие азотистого соединения особенно интересно, поскольку оно отменно работает в бескислородных условиях, а латимерию давно подозревают в том, что она хорошо приспособилась к среде с пониженным содержанием кислорода.
(Из-за этих веществ разлагающийся целакант отвратительно пахнет, с чем пришлось столкнуться мисс Латимер, когда она пыталась сохранить первую пойманную рыбу.)
Спинной мозг заключен в канал, стенки которого укреплены хрящевыми и костными дугами. Каждая дуга переходит в невральный шиповидный отросток, образованный хрящевым стержнем и костной оболочкой. Поскольку хрящ обычно не сохраняется, в ископаемом виде получаем полый шип, который и дал название целакантам (от греч. κοῖλος — «пустой» и ἄκανθα — «колючка»).
Мозговая коробка не цельная, как у других современных костных рыб. Она состоит из двух обособленных монолитных блоков — рыльного, или обонятельно-клиновидного (этмосфеноидного), спереди и затылочно-ушного (отико-окципитального) сзади. Снаружи черепа шов проходит между теменными и заднетеменными костями. Положение очень толстой (4 см в диаметре) хорды, которая прободает задний блок черепа и прирастает вершиной к торцу рыльного, видимо, и предопределило появление межчерепного зазора. Благодаря этому подвижному сочленению пара мощных веретеновидных подчерепных мышц, оттягивая рыло вниз и назад, усиливает захват добычи челюстями. Чтобы задрать его вверх, латимерия подключает удивительный механизм, который сейчас больше ни у кого не встречается. Используя всю мощь плавательной мускулатуры тела, она сжимает хорду; та, будучи несжимаемой, давит прямо в центр переднего блока черепа (нужная дыркато там имеется), толкая его вперед, и пасть разевается.
В рыльном блоке просматривается обширная разветвленная полость, которая в свое время заставила Джона Смита задаться вопросом: не могла ли она вмещать электросенсорный орган, подобный ампулам Лоренцини? Теперь магнитно-резонансная томография подтвердила, что это действительно так. У современных целакантов полость состоит из трех пар взаимно перпендикулярных каналов, создающих трехмерную картину электромагнитного поля и его искажений. Последние подсказывают, что где-то рядом появилась возможная добыча. Расположение полости объясняет загадочное поведение латимерии, которая предпочитает ночами дрейфовать по воле морских течений, время от времени зависая вниз головой. В этот момент ее электролокатор сканирует поверхность дна в поисках незаметной для прочих органов пищи. Интересно, что у палеозойских целакантов электрочувствительные каналы располагались на рыле, как у других рыб, использующих подобное приспособление. Это сходство указывает на то, что и электросенсорный орган, и чувствительность к низкочастотным токам были рыбами унаследованы от общих бесчелюстных предков.
По мере развития мозг латимерии претерпевает значительные изменения. У плода, пока он находится в утробе матери и удивительно похож на головастика, мозговая полость относительно широкая, с короткими обонятельными каналами, уходящими в далеко расставленные носовые капсулы, а ямка для гипофиза — мелкая. Такой же была эта полость у древних костных, хрящевых и пластинокожих рыб. У взрослеющих мальков обонятельные каналы удлиняются, а гипофизарная ямка углубляется вперед и вниз, как у всех мясистолопастных рыб. При этом объем хорды возрастает сильнее, чем объем мозга, который у полутораметровой рыбины оказался буквально вжат в заднюю часть черепа, где занимает всего 1% объема мозговой полости. Остальное пространство заплыло жиром, причем в прямом смысле. Этим целаканты, в том числе латимерия, отличаются от других мясистолопастных рыб и от любых современных позвоночных вообще.
И в кого дальше с таким мозгом можно было эволюционировать? Только в реликты. (Подобный «скукоженный» мозг есть еще у глубоководных акул и скатов.)
Латимерия донесла до нас потрясающее сочетание признаков хрящевых, лучеперых и мясистолопастных рыб, подсказывая, как выглядели их общие предки. Поскольку темпы морфологических преобразований (изменений внешности и внутреннего строения) у целакантов ниже примерно в три раза, чем у костистых рыб, и в шесть раз, чем у четвероногих позвоночных (и важные гены, кодирующие белки, тоже эволюционировали небыстро), латимерию можно считать «живым ископаемым». Этот оксюморон для обозначения подобных форм пришел на ум Чарльзу Дарвину. Однако сказать, что латимерия совсем уж застыла в своей неизменности, тоже нельзя. Даже по сравнению с ее близкой родственницей — меловой макропомой (Macropoma) кое-что стало другим. Например, объемное легкое, скрытое под сплошной скорлупой из крупных костяных пластинок, спаянных известковыми частицами, сократилось до едва заметного «аппендикса», окруженного несколькими мелкими косточками, и погрузилось под мешочек, наполненный жиром. Легкое, открывавшееся в глотку, использовалось для воздушного дыхания и нуждалось в прочной оболочке, чтобы избежать сдавливания. (Впервые необычный костяной орган у ископаемых целакантов описал первооткрыватель динозавров Гидеон Мэнтелл, и ученые долго не могли взять в толк, с чем имеют дело: то ли желудок, как думал Луи Агассис, то ли плавательный пузырь, как считал Артур Смит Вудворд.) Обновленный же орган обеспечивает нейтральную плавучесть на глубине. Плавучести способствуют и отложения жировой ткани в головной и брюшной частях тела, из-за чего даже чучела латимерии сочатся жиром. Наличие легких обычно считается признаком большой продвинутости (мы же всех меряем на свой обезьяний аршин). Однако этот орган был у древних мясистолопастных, а данные по важным генам, влияющим на его развитие, указывают, что легкими могли дышать общие предки всех костных рыб. Причем изначально этот орган был непарным, как на ранних стадиях развития у современных двоякодышащих, латимерии и многопера. Не будем забывать, что концентрация кислорода, растворенного в океане, всю первую половину палеозойской эры еще оставалась низкой, и получать его, заглатывая воздух из атмосферы, было легче. Мясистолопастные и мы, как их потомки, этот архаичный признак сохранили. А костистые нашли более совершенный способ дыхания в насытившейся кислородом водной среде, оставив на память о легких плавательный пузырь, служащий гидростатическим аппаратом. Кроме того, легкие (и плавательный пузырь) играют важную роль в восприятии звуков...
Дальше случилось с целакантами вот что. В мезозойскую эру они изобиловали и в морях, и в пресных водоемах.
Хотя видов было немного, они сильно отличались друг от друга: кто носил шлем на голове, словно пластинокожая рыба, кто отрастил длиннющий мясистый спинной плавник... В полноводных северных реках Африки (там, где сейчас расстилается бескрайняя Сахара), 6‑метровых рыбин охотно вкушали спинозавры. В конце мелового периода «наземные» мясистолопастные исчезли, и остались лишь глубоководные латимерии. К сожалению, о кайнозойских глубоководных отложениях мы можем судить только по материалам морского бурения. Мелкие раковины фораминифер попадаются десятками тысяч, почти незаметные раковинки кокколитофорид — миллионами. Но пробурить скважину и извлечь керн с костями целаканта — это все равно что найти в стоге сена даже не пресловутую иголку, а контактную линзу.
Мезозойские целаканты, в свою очередь, отличались от более древних. Чтобы в этом убедиться, надо копнуть или, поскольку мы имеем дело с рыбами, нырнуть еще глубже — в моря середины девонского периода (410–375 млн лет назад). В ту пору у первых целакантов, подобных шошонии (Shoshonia, названной в честь индейского племени шошонов, населявших американский Вайоминг), широкие грудные плавники несли по краю многочисленные тонкие кожные косточки — плавниковые лучи (лепидотрихии), словно у какихнибудь лучеперых рыб. Но эти косточки, не являясь цельными, состояли из нескольких сочленений. И весь скелет плавников (конечностей) был асимметричным, как у четвероногих. Удлиненные радиалии (по сути, фаланги пальцев) упирались в концы плавниковых лучей, а их, свою очередь, подпирали коротенькие метаптеригии (кости будущего запястья). «Пальцев», конечно, на первых порах получилось многовато, тем более что к каждому метаптеригию подходила пара радиалий, но в девонском периоде в их количестве никто себя не ограничивал. Получилось, что грудной плавник, оставаясь плавательной лопастью, уже приобретал внутренние черты передней лапы.
Гены, регулирующие развитие пояса конечностей вплоть до пальцев (и запястья/щиколотки), которые отсутствуют у всех обычных рыб, как раз «уцелели» у латимерии. Правда, у нее они отвечают за полное развитие плавников и, значит, в ходе эволюции четвероногих переключились на другие детали конечностей. (Это не единственные гены, роднящие целакантов с наземными позвоночными. У них, например, найдено довольно много таких молекулярных элементов, кодирующих рецепторы феромонов и запахов, которые хорошо работают именно в воздушной среде.)
Для нас же сейчас важнее отметить, что ранне- и среднедевонские мясистолопастные рыбы еще сохраняли особенности и лучеперых, и двоякодышащих, и целакантов, но в чем-то уже уподобились первым наземным позвоночным.
Двоякодышащие и предки латимерии как бы остановились на достигнутом, хотя не совсем и не насовсем (и ныне представлены горсткой видов), а лучеперые и будущие четвероноги начали стремительно завоевывать кто океан, а кто сушу (на тот момент — ее узкую прибрежную полосу). Куда эволюция завела первых, мы узнали из предыдущей части книги. А сейчас отправимся в дальний путь вместе со вторыми — шаг за шагом, пусть и шагать им, казалось бы, на первых порах было нечем.
Следует оговориться, что, хотя к современным мясистолопастным рыбам относятся двоякодышащие и кистеперые, ископаемые формы «не пожелали» подчиняться этому удобному для нас разделению на группы. Появлению настоящих четвероногих — с пальцами на лапах — предшествовала длинная череда весьма разных мясистолопастных. В порядке отдаленности родства с четвероногими (и близости к корням родословной) это: элпистостегалии (Elpistostegalia), тристихоптериды (Tristichopteridae), «остеолепидиды» (Osteolepididae), к которым принадлежит гогонас (о нем речь и пойдет ниже), ризодонтиды (Rhizodontida) и поролепиформы (Porolepiformes), включая двоякодышащих. Целаканты с латимерией оказались в самом начале этой многоуровневой развилки. Некоторые из перечисленных названий приходится брать в кавычки, поскольку ныне ученые сомневаются в единстве группы, которую данное название объединяет. Например, целаканты и прочие кистеперые оказались буквально по разные стороны от ветви двоякодышащих, поэтому кистеперые представляют собой несколько неблизкородственных ветвей, и удобное прежде понятие утратило научный смысл.
В австралийском лагерштетте Гого, уже не раз заслуженно упомянутом на страницах этой книги, есть и свой представитель «кистеперых», родовое имя которого, конечно, гогонас (Gogonasus). Изначально эта рыба описана всего по нескольким косточкам рыла, но в конце концов удалось найти целый череп и, значит, понять, как были устроены мозговая и другие полости (рис. 10.1). Скажем, по наличию заметных затылочных вырезок-углублений, где располагалось парное брызгальце, череп гогонаса резко отличается от такового латимерии. Однако он весьма похож на эту часть скелета у их родственников, более продвинутых в сторону четвероногих, — пандерихта и тиктаалика. Последние использовали пару первых жаберных щелей, сильно преобразованных и обращенных вверх, чтобы с шумом вдыхать воздух. (А их потомки, у которых эти вырезки станут ушными, — и чтобы слышать.)
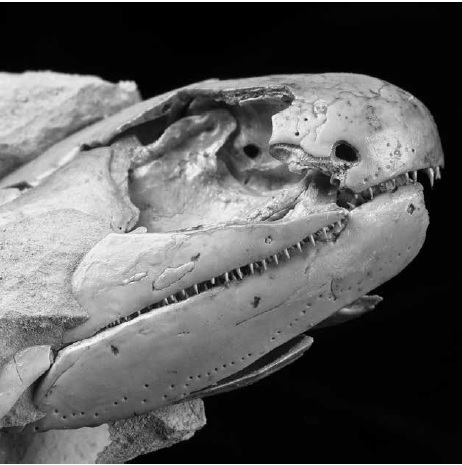
Механизм дыхания через брызгальца изучен у современных многоперов. Сначала рыба выдавливает, сжимая тело, воздух из легкого в ротоглоточную полость, которая расширяется, и газ покидает ее сквозь обычные жаберные щели. Полость (весьма обширная у гогонаса), как следствие, сокращается, и через открывшийся клапан брызгальца туда устремляется свежий воздух, вновь ее расширяя. Клапан закрывается, полость стискивается, легкое наполняется. (Конечно, делать все это надо с закрытым ртом.) Гогонас мог дышать и через ноздри: тогда воздух в полость рта поступал через хоаны — парные отверстия в переднем внутреннем отделе черепа, ограниченные предчелюстными, верхнечелюстными и нёбной костями и сошником. Благодаря описанному «нехитрому» механизму рыбе не требовалось открывать рот при каждом вдохе и выдохе.
Вероятно, с особенностями «дыхательно-слухового » комплекса было связано еще одно преобразование скелета: гиомандибула укоротилась и из вертикального положения перешла в почти горизонтальное, заняв место рядом с брызгальцем. Этой части прежней жаберной дуги оставалось совсем чуть-чуть, чтобы обратиться в первый элемент нашего слухового аппарата — стремечко. Без дела эта косточка даже в таком положении не пребывала и с помощью оперкулярного отростка помогала раскрывать широкие и тяжеловатые жаберные крышки.
Некоторые черты сближали гогонаса и с целакантами. Например, полость в рыльном блоке черепа, к которой подходил канал поверхностной ветви зрительного нерва, как у латимерии. У последней в такой же полости находится электрочувствительный орган.
Сам мозг гогонаса был намного крупнее, чем у целакантов, с тремя развитыми полукружными каналами лабиринта внутреннего уха и «широко раскрытым» третьим глазом.
Плечевой пояс гогонаса включал ключицы, задневисочные кости и несколько покровных элементов. Это были (от спины к брюху): супраклейтрум, аноклейтрум и клейтрум — каждая кость, конечно, в двух версиях, правой и левой. Вместе с ключицами они образовывали как бы квадратный обруч позади черепа. Изнутри к клейтруму подходил скапулокоракоид — это уже кость внутреннего скелета (окостеневший хрящ), будущие лопатка и коракоид лягушек, вместе взятые. Задневисочные кости намертво сочленяли плечевой пояс с черепом, ключицы примыкали к нижним элементам жаберных крышек. (Вероятно, имелась и межключичная кость, замыкавшая обруч снизу, но она не сохранилась.) Судя по следам мышечных креплений к отросткам костей, плечевой пояс уже двигался, но независимость от головного отдела еще не приобрел.
Скелет самой передней конечности более узнаваем: плечевая кость, за которой следовали локтевая и лучевая. Правда, отходила плечевая кость от скапулокоракоида, все кости были плоскими, а лучевая — шире и длиннее, чем локтевая. К ним примыкали кости запястья: ульнаре, или локтезапястная, и промежуточная. По-настоящему рыбьими чертами гогонаса были ромбовидная космоидная чешуя и покровные кости черепа сходного строения. (Что значит «космин», нетрудно догадаться по однокоренному слову «косметика», образованному от греч. κοσμέω — «украшать».) Такие элементы украшены тонкой блестящей пористой эмалью, под которой расположен дентиновый слой, весь пронизанный многочисленными колбовидными каналами и пульпарными полостями с расходящимися от них пучками канальцев. Глубже следуют кость и пластичный эласмодин. (Такие покровы встречаются только у ископаемых мясистолопастных, но для чего служили разнообразные полости, пока неизвестно.)
Были у гогонаса и межчерепной сустав, хотя, видимо, не столь подвижный, как у латимерии, и развитые жаберные крышки (из трех костей каждая). Так что он еще представлял собой рыбу с чешуей, расходившимися веером плавниковыми лучами и объемной хордой, но многие детали его скелета, в первую очередь конечностей и ушной части черепа, предвосхитили строение настоящих четвероногих. В совокупности особенности гогонаса, даже электрорецепция, позволяют предположить, что он был маневренным мелководным существом, способным в случае необходимости (застрял в луже во время отлива) глотнуть свежего воздуха. Среди рыб, похожих на четвероногих, гогонас приходился одним из ближайших родственников четвероногим, похожим на рыб.
Читайте также
Исследователи определили, когда появились первые четвероногие
Девонский период стал переломным моментом для мясистолопастных: в то время они были гораздо разнообразнее лучеперых. Но, главное, за несколько десятков миллионов лет рыбы, напоминавшие современную латимерию, превратились в расхаживающих по суше животных. Движение было не быстрым, но поступательным. И снова — рыба эволюционирует с головы: строение головного отдела изменялось в два раза быстрее, чем прочие особенности скелета. Прогресс не остановило даже массовое вымирание, случившееся в середине позднедевонской эпохи (372 млн лет назад). Скорее наоборот: разнообразие мясистолопастных рыб пошло на спад, а четвероногие пережили бурный расцвет. Эволюция не искала магистральных путей и, кроме непосредственных предков современных и древних «лягушек», мы видим огромное разнообразие животных, которые в устройстве отдельных органов на тот момент далеко обогнали настоящих четвероногов. Кому-то необходимо было лишь собрать все наработки в едином теле...
В 1952 г., когда Смит разыскал вторую особь целаканта и «представил» ее в Кейптауне премьер-министру тогдашнего Южно-Африканского Союза Даниелю Малану (создателю системы апартеида и, как все религиозные диктаторы, ярому борцу с любым свободомыслием), тот поинтересовался: «Вы хотите сказать, что и мы некогда выглядели таким образом?» На что ученый ответил: «Я встречал людей и пострашнее».